Alfred Lang | University of Bern, Switzerland |  |
Edited Book Chapter 1973 |
Der Umgang mit Dauer: Ein neues Modell der inneren Uhr [1] | 1973.01 @Time @Perc @Act |
24 / 42 KB + 3 figs., 1 tabl.
Last revised 98.10.31 |
Pp. 587-596 in: REINERT, G. (Ed.) 1973. Bericht über den 27. Kongress der Deutschen Gesellschaft für Psychologie in Kiel 1970. Göttingen: Hogrefe. | © 1998 by Alfred Lang |
info@langpapers.org |
Scientific and educational use permitted |
Home || |
Inhalt
Zusammenfassung. Nach einer kurzen Kritik am
mechanistischen Einheitenzählmodell (Momenttheorie) wird ein
Alternativmodell der inneren Uhr als ein multiples System skizziert.
Danach ist die Zeit des Individuums das organisierte Insgesamt der
zentralen Repräsentation von vielen, ein breites
Frequenzspektrum deckenden und partiell autonomen biologischen
Rhythmen. Die wahrgenommene oder hergestellte Dauer eines gegebenen
Ereignisses wird als eine Funktion der Kongruenz zwischen der
Repräsentation des Ereignisses mit mehreren Teilprozessen des
Systems bei unterschiedlicher Gewichtung derselben
aufgefaßt.
Abstract. After a brief criticism of the mechanistic unit
counting model (moment theory) the author Sketches a new model of the
inner clock, suggesting the idea of a multiple system. Accordingly,
the time of the individual is the organized sum total of the central
representation of many and partially autonomous biological rhythms
which cover a broad frequency spectrum. The perceived or produced
duration of a given event is assumed to be a function of the
congruence between the central representation of the event and
several, differentially weighted part-processes of the system.
Top of Page
Nach den
Einheitenzählmodellen sollen Wahrnehmung, Schätzung,
Herstellung und Vergleich von Intervallen bestimmter Dauer dadurch
möglich sein, daß wir über ein Einheitsintervall und
einen Zählmechanismus, d. h. eine innere Uhr analog den
physikalischen Uhren, verfügen. Dieses Prinzip geht auf
Vorschläge von COLERIDGE (1817), VON BAER (1864), VON
UEXKÜLL (1928) und anderen zurück und wurde besonders von
STROUD (1955, 1967) in Form der kybernetischen Momenttheorie und von
CREELMAN (1962) in einer stochastischen Variante erneuert (vgl. auch
LANG 1971 a). Einen Verifikationserfolg kann man weder den
älteren (z. B. MÜNSTERBERG 1889; BRECHER 1932) noch den
neueren (z. B. TREISMAN 1963; MICHON 1967) empirischen
Bemühungen attestieren.
Die Hauptschwierigkeit der Einheitenzählmodelle liegt darin,
daß es nicht gelingt, die Einheitsdauer (den Moment oder die
gezählten Ereignisse) und das Zählprinzip voneinander
unabhängig in den Griff zu bekommen. Das ist aber notwendig,
weil das Einheitenzählmodell multiplikativ ist. Die
wahrgenommene oder produzierte Dauer ist gedacht als ein Produkt aus
der Erstreckung der Einheitsdauer d(m) und der Menge oder Anzahl z
der solcherart abgezählten Einheiten
D = f [z d(m)]Die verfügbare Forschung zeigt nun hinreichend, daß
weder die Einheitsdauer eine konstante, empirisch faßbare
Größe, noch daß der Zähler ein einfacher
additiver Akkumulator ist (vgl. FRASSE 19672; BERGIUS
1969). Die Untersuchung des Moments setzt damit spezifische Annahmen
über den Zählmechanismus voraus und umgekehrt, so daß
sich die Forschung im Zirkelschluß bewegt. Soweit ich die
Literatur überblicke, gibt es zur Frage des Zählers
ausschließlich Spekulation; die vorgeschlagenen
Operationalisierungen des Moments sind problematisch, und von keiner
von ihnen ist nachgewiesen, daß sie für Zeitwahrnehmung
und Zeitverhalten in eindeutiger Weise relevant ist.
Top of Page
Die "innere Uhr"
als multiples System
Das Modell einer multiplen inneren Uhr geht von der Annahme aus,
daß im lebenden Organismus jederzeit eine Vielzahl von zeitlich
strukturierten Vorgängen, und zwar in der Mehrzahl zyklischen
und quasizyklischen Prozessen, abläuft. Diese Vorgänge
werden wahrscheinlich nicht wie im Computer von einem einzigen
zentralen Taktgeber ausgelöst oder gesteuert, sondern sie sind
voneinander wenigstens teilweise unabhängig. Ober die
anatomisch-physiologische Grundlage dieser Prozesse mache ich mir
zunächst keine spezifischen Vorstellungen. Es genügt mir
die Tatsache, daß Biologie und Medizin eine große Zahl
solcher "biologischer Rhythmen" nachgewiesen haben.
Frequenzmäßig sind diese Rhythmen über ein sehr
breites Spektrum verteilt; man findet Rhythmen mit Periodenlänge
in Größenordnung von Sekundenbruchteilen bis zur
Größenordnung von Monaten, ja vielleicht Jahren (vgl. z.
B. SOLLBERGER 1965). Immerhin scheint festzustehen, daß die
Mehrzahl der bisher isolierten Rhythmen endogener Natur ist. Nicht
beliebige Frequenzen kommen vor, sondern bei einem gegebenen
Prozeßtypus stets ganz bestimmte, die Art und das Individuum
kennzeichnende Werte. Die Rhythmen sind ferner zumindest partiell
autonom, insofern sie der Beeinflussung durch andere Rhythmen sowie
der Synchronisation in Frequenz und Phase an periodische
Umweltereignisse zwar unterworfen, aber nicht beliebig, sondern stets
nur in begrenztem Ausmaß unterworfen sind. Ich erinnere zum
Zweck der Konkretisierung dieses Begriffes der partiellen Autonomie
an die bekannten Untersuchungen VON HOLSTS (1937) über die
relative Koordination der Extremitätenbewegungen bei Fischen und
Säugern.
Bei Pflanzen und niederen Tieren sind nun im allgemeinen die
biologischen Rhythmen mit je bestimmten Auslösemechanismen
verbunden, und jeder Rhythmus wird stets nur in seinem
zugehörigen Funktionszusammenhang wirksam. Bei höher
organisierten Lebewesen ist wohl ein großer Teil aller Rhythmen
im Hinblick auf ihre Koordination zu einem funktionsgerechten
Handlungsinsgesamt im Zentralnervensystem repräsentiert. Damit
verbunden ist eine Einschränkung der Spezifität der
Rhythmen. Das gilt auf kortikalem Niveau (man kann ja
willkürlich im Rhythmus des Atmens schreiten oder kauen usw.),
wahrscheinlich aber auch in allgemeinerer Weise; zum Beispiel
beeinflußt der circadiane Aktivitätszyklus ganz
verschiedenartige Effektorrhythmen (MILLS 1966). Mit anderen Worten:
Die Periodik eines gegebenen Rhythmus kann für einen andern
Prozeß, welcher nicht primär demselben
Funktionszusammenhang angehört, relevant werden.
Damit habe ich einen der Wege skizziert, die mich zur Formulierung
der folgenden allgemeinen Hypothese geführt haben (vgl. auch
LANG 1970): Die Zeit des Individuums ist das organisierte Insgesamt
der zentralen Repräsentation von vielen, ein breites
Frequenzspektrum deckenden und partiell autonomen biologischen
Rhythmen. Infolge der relativen Unspezifität der zentralen
Repräsentation der biologischen Rhythmen ist denkbar, daß
fast beliebige Ereignisse der wahrgenommenen Umwelt und Innenwelt auf
diese multiple innere Uhr abgebildet (Zeitwahrnehmung) bzw. daß
diese multiple innere Uhr zur Herstellung von fast beliebigen
zeitlich strukturierten Akten (Zeitverhalten) eingesetzt werden kann.
Die Dauer eines gegebenen Ereignisses ist demnach eine Funktion der
Kongruenz zwischen der Repräsentation des Ereignisses mit
mehreren Teilprozessen der multiplen Uhr:
D = f [z1 d(m1),
z2 1 d(m2), . . . , zn
d(mn), . . . ]Dabei stelle ich mir nun vor, daß je nach Art und
Erstreckung des zu schätzenden Ereignisses diesem oder jenem der
verschiedenen Rhythmen ein unterschiedliches Gewicht zukommt.
Ich beschränke mich darauf, zwei wichtige Konsequenzen des
multiplen Modells der inneren Uhr zu nennen: (1) Die multiple Uhr ist
in hohem Maß stabil gegen Störungen. Das entspricht der
tatsächlichen Alltagsleistung unseres Zeitverhaltens im Schlaf
und im kognitiv unbelasteten Wachzustand, während
möglicherweise die Standardexperimente zur Zeitwahrnehmung mit
ihren so widersprüchlichen Ergebnissen im Sinne einer
Partialisierung der multiplen Uhr wirken, so daß durch die
Dominanz eines einzelnen und den weitgehenden Wegfall der
übrigen Teilprozesse eine analoge Situation entsteht wie beim
Größenkonstanzverlust unter Wegfall der unabhängigen
Distanzinformation. (2) Die multiple Uhr ist ökonomisch, da zur
Schätzung der Dauer eines gegebenen Ereignisses stets auch
Teilprozesse mit Periodenlängen von ähnlicher
Größenordnung eingesetzt werden. Beim einfachen
Einheitenzählmodell ist hingegen die für kurze Ereignisse
notwendige hohe Präzision bei längeren Ereignissen reine
Verschwendung; es muß dann postuliert werden (z. B. TREISMAN
1963), daß sie in der Verarbeitung irgendwo verlorengeht, da
bekanntlich die Unterschiedsempfindlichkeit mit zunehmender Dauer
eher abnimmt.
Top of Page
Experimentelle
Strategie und Versuchsanordnung
Das Prinzip meines Experimentierens besteht darin, die
wechselseitige Abhängigkeit von ineinander verschachtelten
Dauerschätzungen festzustellen. Dabei geht es mir zunächst
um den Nachweis, daß ineinander verschachtelte
Dauerschätzungen eines Individuums Systemcharakter haben. In
zweiter Linie möchte ich allmählich dazu kommen, einzelne
Teilprozesse des Systems im oben beschriebenen Sinn zu isolieren und
derart zu manipulieren, daß ihre Rolle für das
Zeitverhalten insgesamt spezifiziert werden kann. Ob diese
Bemühungen schließlich zu einem falsifizierbaren Modell
der multiplen inneren Uhr führen werden, das auch in formaler
Hinsicht die heutigen Ansprüche zu befriedigen vermag,
müssen Zeit und Geduld erweisen. Aus erwähnten Gründen
experimentiere ich grundsätzlich innerhalb des Individuums; die
allgemeinere Gültigkeit der Aussagen wird durch Aufzeigen
analoger Strukturen bei mehreren Individuen gerechtfertigt.
Die Vpn -- mehrheitlich Hausfrauen, die während mehreren
Wochen täglich eine l- bis 2-stündige Sitzung absolvieren
-- bekommen also den Auftrag, durch Knopfdruck ein bestimmtes
Intervall, z. B. 1 bzw. n Minuten zu produzieren. Ein paar
Rückmeldungen über die Leistung zu Beginn jeder Sitzung
vermindern ziemlich rasch die Varianz dieser längeren
Produktionen sowie das anfängliche Unsicherheitsgefühl,
ohne physische Uhr zu arbeiten. Während jeder Sitzung produziert
nun die Vp wiederholt dasselbe längere Intervall ohne
Rückmeldung. Das Intervall wird ausgefüllt durch ein
zeitlich strukturiertes Verhalten, welches die Beurteilung eines oder
mehrerer Teilprozesse der multiplen Uhr bzw. Aussagen über die
Periodik von m, erlauben soll. Dabei wird angenommen, daß die
Zeitpunkte der einzelnen Akte dieses zeitlich strukturierten
Verhaltens auf irgendeine Art und Weise Manifestationen der
postulierten Teilprozesse der multiplen Uhr darstellen. Ich muß
also hier eine methodisch bedingte Zusatzannahme einführen,
über deren Berechtigung erst post hoc entschieden werden kann.
Die Verifikation des Modells bestätigt die Richtigkeit der
Zusatzannahme; hingegen ist bei seiner Falsifizierung unsicher, ob
das Modell als Ganzes oder nur diese Zusatzannahme ungültig ist.
Immerhin ist traditionellerweise die Spontanfrequenz von Tapping als
ein Indikator des persönlichen Tempos oder desgleichen
betrachtet worden, und diese stellt natürlich in meinem Ansatz
den Fall dar, wo ein Teilprozeß aufs Mal manifestiert wird. Die
Manifestation mehrerer Teilprozesse zugleich wird dadurch erreicht,
daß die Vpn irgendwelche variierende Aspekte von dargebotenen
Bildern in Betrachtungsdauer kodieren. Praktisch sieht das so aus,
daß zu Beginn der längeren Produktion ein Bild erscheint,
z.B. 3 längliche Rechtecke (mmmm mmmm mmmm ). Jedesmal, wenn die
Vp mit einem Griffel ein Plättchen berührt, wechselt
unmittelbar das Bild: Es erscheinen jedesmal Rechtecke von
unterschiedlicher Länge (z. B. mm mm mm oder m m m m m m). Die
Vp hat nun den Auftrag, den Bildwechsel zeitlich so zu strukturieren,
daß die Betrachtungsdauer jedes einzelnen Bildes ungefähr
proportional zur Länge der Rechtecke auf dem betreffenden Bild
ist. Das heißt: Je kürzer die Rechtecke, desto kürzer
die Betrachtungsdauer, und je länger die Rechtecke, desto
länger die Betrachtungsdauer; natürlich werden auch der
umgekehrte Kodierschlüssel sowie auch andersartiges Bildmaterial
eingesetzt. In der Wahl der Dauer der einzelnen
Betrachtungsintervalle ist die Vp völlig unbeeinflußt und
frei mit Ausnahme der Restriktion, daß gleiche
Strichlängen nach Möglichkeit in stets gleiche
Betrachtungsdauern kodiert werden sollen, und zwar ohne jedes
Zählen oder Taktieren. Die meisten Vpn lernen und stabilisieren
diese Kodiertätigkeit im Verlauf von 2 bis 5 Sitzungen zu einer
erstaunlichen Regelmäßigkeit. Trotz der offensichtlichen
Monotonie empfinden praktisch alle Vpn diese Tätigkeit als eine
anspruchsvolle und vor allem faszinierende Aufgabe.
Die Bilderreihen werden elektronisch erzeugt und auf einem
Oszillographen dargeboten. In einer Variante kommen auch Dia-Serien
zur Verwendung. Pro Versuchseinheit, d. h. im Zeitraum der von der Vp
produzierten n Minuten, werden gewöhnlich zwei bis vier
verschiedene Bilder oder Bildklassen mit einer irrelevanten Variation
innerhalb der Klassen in zufälliger Reihenfolge und mit
bestimmten Wahrscheinlichkeiten dargeboten. Die physische Dauer jedes
einzelnen Betrachtungsintervalls wird elektronisch gemessen. Ein
kleiner Rechner erlaubt die unmittelbare Verarbeitung, Speicherung
und Darstellung dieses zeitlich komplex strukturierten Verhaltens in
Form von Häufigkeitshistogrammen über der
Betrachtungsdauer. Pro Versuchseinheit fallen als Ergebnis jeweils
ein Häufigkeitshistogramm der Betrachtungsdauern sowie ein Wert
für die produzierte Dauer des längeren Intervalls an.
Letzteres wird zum Zweck der Vergleichbarmachung als Quotient
(Produktion durch Standard) verrechnet und einer logarithmischen
Transformation unterzogen; die resultierenden u-Werte zeigen bei 500
die Obereinstimmung der inneren mit der sozial-physikalischen Uhr an;
u < 500 bedeutet Unterproduktion, z.B. entspricht u = 403 einer
Produktion von 80% des Standards; u > 500 bedeutet Oberproduktion,
z. B. u = 579 entspricht 140%.
Top of Page
Einige Ergebnisse
und ihre Bedeutung
In der geschilderten Versuchsanordnung sind sicher eine Reihe von
Faktoren wirksam, die ich experimentell nicht (oder noch nicht)
beherrsche. Die relativ großen Streuungen nötigen, jeweils
über eine gewisse Zahl von Versuchseinheiten Durchschnitte zu
bilden. Innerhalb dieser Klassen, die in der Regel aus Daten
desselben oder wenigstens unmittelbar aufeinander folgender
Versuchstage einer Vp gebildet sind, ist eine Spontanvariation des
Zeitverhaltens gegeben, welche sich einerseits als Prüfstreuung
eignet (und zwar als recht konservative Prüfstreuung, die nur
deutliche Effekte akzeptiert), und welche anderseits auch zur
Hypothesenbildung für das weitere Experimentieren herangezogen
werden kann.
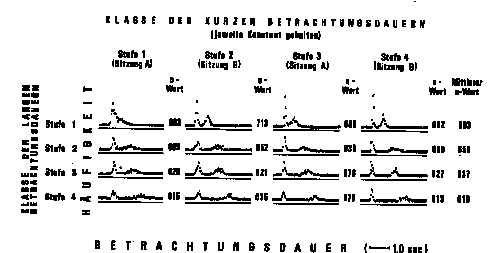
Abbildung 1. Häufigkeitsdiagramme und durchschnittliche
u-Werte der Versuchsperson FJ in einem 4x4Versuchsplan mit zwei
Betrachtungsdauer-Klassen. Die Sitzung B erfolgte am Tage nach der
Sitzung A um dieselbe Uhrzeit. Der Standard der längeren
Produktion betrug eine Minute. Jedes einzelne der 16 Histogramme ist
die Summe, und der zugehörige u-Wert ist der Durchschnitt von
vier Versuchseinheiten. Je höher die Punkte über der
jeweiligen Grundlinie liegen, desto mehr Betrachtungsintervalle der
betreffenden Dauer sind von der Versuchsperson hergestellt worden.
Eine doppelte Varianzanalyse ergibt, bei nichtsignifikanter
Spaltenwirkung und Spalten x ZeilenWechselwirkung, auf dem 5%-Niveau
gesicherte Unterschiede zwischen den Zeilen (F = 3,28 bei 3 und 57
Freiheitsgraden). Die Zeilenunterschiede erklären 16,6 Prozent
der erfaßten Varianz.
Die Abbildung 1 enthält die Ergebnisse von insgesamt 64
Versuchseinheiten einer Vp. Durch ein entsprechendes Angebot von
Rechtecklängen wurde die Vp zur Herstellung von 2 Klassen von
Betrachtungsintervallen, nämlich von relativ kürzerer und
von relativ längerer Dauer, induziert. In jeder Spalte sind die
kürzeren Betrachtungsdauern konstant gehalten (die linken
Verteilungsberge, die jeweils ziemlich genau übereinander
liegen), während die längeren Betrachtungsdauern in 4
Stufen manipuliert worden sind. Mit zunehmender Dauer des
längeren Betrachtungsintervalls (von oben nach unten) nimmt die
subjektive Dauer der produzierten Minute sukzessive ab (u-Werte).
Dieses Ergebnis widerspricht natürlich der naiven Erwartung,
daß längere Einheitsdauern zu längeren
Dauerschätzungen führen müßten. Ebenfalls im
Widerspruch zur Voraussage, welche die Theorie von FRAISSE (19672)
machen würde -- je dichter die Ereignisse, desto schneller die
innere Uhr, desto kürzer die Produktion --, tritt also bei der
Herstellung des längeren Intervalls eine Kompensation für
die länger werdenden Betrachtungsintervalle ein, und zwar eine
Überkompensation.
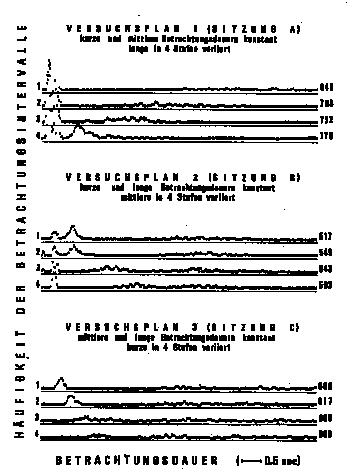
Abbildung 2. Häufigkeitshistogramme und durchschnittliche
u-Werte der Versuchsperson FK in drei Versuchsplänen mit je drei
Betrachtungsdauer-Klassen. Die Sitzungen A, B und C erfolgten an drei
verschiedenen Tagen. Die Zahl der Versuchseinheiten pro Histogramm
beträgt 5 im Versuchsplan 1, 9 im Versuchsplan 2 und 7 im
Versuchsplan 3. Die Prüfung der Unterschiede zwischen den
u-Werten erfolgte in separaten einfachen Varianzanalysen (siehe
Text).
Tabelle 1 Signifikanzniveau der Mittelwertunterschiede (SN) und
Prozentsatz der Varianz (PV) in den u-Werten, die durch
unterschiedliches Reizangebot bei 3 Standardlängen der
Produktion erklärt werden (Vp : FK)
Standard | Variation der Betrachtungsdauera |
|---|
kurz | mittel | lang |
|---|
SN | PV | SN | PV | SN | PV |
|---|
1/2 Minute | n.s. | 0% | -- | -- | s.s.b | 36% |
|---|
1 Minute | n.s. | 1% | n.s. | 0% | n.s. | 48% |
|---|
| n.s. | 11% | n.s. | 9% | s.c | 38% |
|---|
| | | | | s.s.b | 58% |
|---|
2 Minuten | -- | -- | -- | -- | n.s. | 21% |
|---|
- a Bei Konstanthaltung der beiden übrigen Betrachtungsdauern.
- b p < .0l.
- c p < .05.
Auf dem Hintergrund des oben skizzierten multiplen Modells der
inneren Uhr würde dies bedeuten, daß hier zwei
verschiedene Teilprozesse manifest geworden sind, von denen sich nur
einer für Dauerschätzungen von 1 Minute als relevant
erweist, und zwar in einer Art und Weise, die den Schluß auf
den Systemcharakter des Gesamtprozesses nahelegt. In Abbildung 2 sind
Ergebnisse einer andern Vp zusammengefaßt, wobei von 3 Klassen
von Betrachtungsdauern jeweils 2 konstant gehalten und die dritte in
4 Stufen variiert worden ist. Während in den beiden unteren
Versuchsplänen die Variation von kurzen und mittleren
Betrachtungsdauern ohne Einfluß auf die Dauer der längeren
Produktion bleibt, ergibt im obersten Versuchsplan die Variation der
langen Betrachtungsdauern eine deutliche systematische Variation der
u-Werte, die den oben dargestellten Resultaten entspricht. Tabelle 1
gibt weitere Resultate derselben Vp im Überblick. Von 3
manifestierten Teilprozessen der multiplen Uhr scheint demnach nur
der längste für Dauerschätzungen in der
Größenordnung von Minuten relevant zu sein.
Der Anspruch auf allgemeinere Gültigkeit dieser Aussagen sei
durch die in Abbildung 3 dargestellten Ergebnisse dreier weiterer Vpn
bekräftigt. Die Histogramme (in logarithmischer Ordinatenskala)
zeigen deutlich, daß jede Vp trotz des gleichen Reizangebotes
ganz unterschiedliche Dauern der kurzen, mittleren und langen
Betrachtungsintervalle wählt. In jedem Fall nehmen jedoch mit
der Verlagerung des Zeitverhaltens auf längere
Betrachtungsdauern die u-Werte ab.
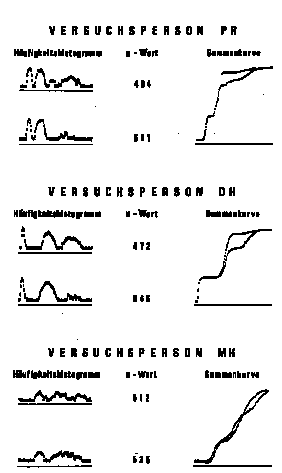
Abbildung 3. Häufigkeitshistogramme, durchschnittliche
u-Werte und Summenkurven von drei Versuchspersonen mit drei
Betrachtungsdauer-Klassen. Die Manipulation des längsten
Betrachtungsintervalles erfolgte hier durch ein unterschiedliches
Angebot an Diapositiven, wobei die Dichte von zufälligen
Lichtpunktanordnungen in Betrachtungsdauern zu kodieren war. Bei
allen Versuchspersonen war im oberen Histogramm gegenüber dem
unteren ein Teil der Dias mit mittlerer Dichte durch solche mit
höherer Dichte ersetzt worden. Die statistische Beurteilung der
Unterschiede in den u-Werten ist hier wegen der
erwartungsgernäss signifikanten Interaktion Versuchspersonen X
Reizstufen problematisch.
In den kumulativen Summenkurven wird noch deutlicher die jeweilige
Übereinstimmung bei den kurzen und die relative Verlagerung von
den mittleren auf die langen Betrachtungsdauern erkennbar. Bei Vp MH
kommt es zu einer Art Kontrastbildung im Gesamtsystem, insofern hier
die Vermehrung der längeren Betrachtungsdauern eine relative
Verkürzung der mittleren zur Folge hat.
Schlussbemerkung
Wenn ich die gesamten bisher vorliegenden Ergebnisse von 10 Vpn
vorsichtig zusammenfassend zu interpretieren wage, dann lassen die
demonstrierten Abhängigkeiten die folgenden Verallgemeinerungen
zu: (1) Die Verteilungen der Betrachtungsdauern haben
Systemcharakter, d. h. es werden nicht beliebige, sondern bevorzugt
Intervalle von bestimmter Dauer hergestellt, und häufig zieht
die Veränderung eines Intervalls Verschiebungen in andern nach
sich. (2) Die Betrachtungsdauerverteilungen sind in systematischer
Weise für das Zeitverhalten relevant, insbesondere für
Schätzungen in der Größenordnung von wenigen
Minuten.
Es dürfte sich also wohl lohnen, durch parametrische
Untersuchungen, verbunden mit einer vorsichtigen Mathematisierung,
die Vorstellung der multiplen inneren Uhr in ein präziseres
Modell umzusetzen. Ein Ansatz zu einer echten Alternative zum
mechanistischen Einheitenzählmodell liegt hier vor, welcher
vielleicht formal nicht so leicht zu bewältigen sein wird,
welcher jedoch viel besser zu unserem allgemeinen Wissen vom
Wahrnehmen und vom Handeln paßt.
Top of Page
Note
[1] Mit Unterstützung des
Schweizerischen Nationalfonds zur Förderung der
wissenschaftlichen Forschung, Projekt Nr. 5247.1. <<<<<
Literatur
BERGIUS, R. 1969. Vom Zeitsinn zum Verhaltensparameter Zeit. In:
IRLE, M. (Ed.) Bericht über den 26. Kongreß der Deutschen
Gesellschaft für Psychologie Tübingen 1968. Göttingen:
Hogrefe. p. 1-21.
BRECHER, G. A. 1932. Die Entstehung und biologische Bedeutung der
subjektiven Zeiteinheit des Moments. Zeitschrift für
Vergleichende Physiologie 18, 204-243.
COLERIDGE, T. S. 1817. Biographia Literaria (Republikation: 1908.
London: Dent).
CREELMAN, C. D. 1962. Human discrimination of auditory duration.
Journal of the Acoustical Society of America 34, 582-593.
FRAISSE, P. 1967, 2. Aufl. Psychologie du temps. Paris: Presses
Universitaires de France.
LANG, A. 1970. Ober den Primat der subjektiven
Wahrnehmungsdimension, dargestellt am Beispiel der Zeitpsychologie.
Schweizerische Zeitschrift für Psychologie und ihre Anwendungen
29, 45-51.
LANG, A. 1971 (a). Die innere Uhr als multiples System: Beitrag
zur Psychologie der Zeitwahrnehmung und des Zeitverhaltens. Bern:
Psychologisches Institut der Universität Bern
(Unveröffentlichtes Manuskript).
LANG, A. 1971 (b). Psychologie der Zeit. In: ARNOLD, W., EYSENCK;
H. J. 8r MEILI, R. (Ed.) Lexikon der Psychologie. Band 1. Freiburg i.
Br.: Herder. p. 38-43.
MICHON, J. A. 1967. Timing in temporal tracking. Soesterberg
(Niederlande): Institute for Perception RVO-TNO.
MILLS, J. N. 1966. Human circadian rhythms. Physiological Review
46, 128-171.
MÜNSTERBERG, H. 1889. Beiträge zur experimentellen
Psychologie, Heft 2. Freiburg i. Br.: Siebeck.
SOLLBERGER, A. 1965. Biological rhythm research. New York:
Elsevier.
STROUD, J. M. 1955. The fine structure of psychological time. In:
QUASTLER, H. (Ed.) Information theory in psychology. Glencoe, Ill.:
Free Press. p. 174-207.
STROUD, J. M. 1967. The fine structure of psychological time.
Annals of the New York Academy of Science 138, 623-631.
TREISMAN, M. 1963. Temporal discrimination and the indifferente
interval: Implications for a model of the "internal clock".
Psychological Monographs 77 (13, Whole No. 576).
VON BAER, K. E. 1864. Welche Auffassung der lebenden Natur ist die
richtige? In: VON BAER, K. E. (Ed.) Reden, gehalten in
wissenschafflichen Versammlungen, und kleinere Aufsätze
vermischten Inhalts. 1. Teil: Reden. Rede No. 5, gesprochen 1860. St.
Petersburg: Schmitzdorff. p. 237-283.
VON HOLST, E. 1937. Vom Wesen der Ordnung im Zentralnervensystem.
Naturwissenschaften 25, 625-631, 641-647.
VON UEXKÜLL, J. 1928, 2 Aufl.. Theoretische Biologie. Berlin:
Springer.
Top of Page